
细胞的分类
原核细胞
原核细胞:一类没有成型细胞核的细胞,其细胞核为拟核。纽荷尔细胞显微镜使用方法,其 DNA 不与蛋白 质结合,在细胞里盘曲折叠,没有膜包裹。仅含有核糖体。一般以裂殖方式 增殖。主要有:细菌、蓝藻、放线菌、支原体和衣原体等。真核细胞:一类具有完整细胞核的细胞。其 DNA 与蛋白质结合,有膜包 裹。有多种特定功能的细胞器。细胞增殖的方式除了无丝分裂方式外,还有 有丝分裂和减数分裂(形成配子)。主要有:动植物、酵母菌等。古细菌:有时也称为“第三类生物”,原来曾归入原核生物的细菌域,现在已经分出。往往生存在其它两域生物无法生存的极端环境中。具有原核 生物的某些特征,如无核膜及内膜系统;也有真核生物的特征,如以甲硫氨 酸起始蛋白质的合成、核糖体对氯霉素不敏感、聚合酶和真核细胞的相似、RNA DNA 具有内含子并结合组蛋白; 此外还具有既不同于原核细胞也不同于真核细 胞的特征。

第二篇:细胞死亡的分类题库
细胞死亡是多细胞生物生命过程中重要的生理或病理现象。细胞死亡有很多种,基于不同的分类标准,其分类结果也不一样。细胞死亡有程序性和非程序性之分,后者即坏死。程序性细胞死亡按其发生机制不同可以分为凋亡、自吞噬性程序性细胞死亡、Paraptosis、细胞有丝分裂灾难、胀亡等。也有很多学者按照死亡时细胞的形态特征进行分类:按照细胞核形态可以分为凋亡、凋亡样程序性细胞死亡、坏死样程序性细胞死亡和坏死;按照Clarke形态学分类,程序性细胞死亡分为I、II、III类。形态学分类与机制分类有很大的重叠,我们总结了它们的对应关系。
一、前言
多细胞生物的发育及生存依赖于其细胞分裂增殖和死亡之间的平衡,一旦这种平衡被打破,就会发生胚胎发育异常、退行性疾病以及癌症等。所以在进化过程中,多细胞生物逐渐拥有了复杂而精密的调节机制维持这种平衡。
细胞的死亡形式多种多样,在过去的150年其分类主要基于形态学。而在最近的30年里,由于死亡分子机制方面的研究取得长足进步,使得死亡的分类更加科学化。但是,由于认识的局限性,细胞死亡分类的现状是形态和机制并存,甚至给人一种混乱的感觉,本文将对现有的细胞死亡分类加以综述,并尽量将形态学分类与机制分类联系起来。
二、非程序性细胞死亡
程序性细胞死亡的共同点在于它们是细胞主动的死亡过程,能够被细胞信号转导的抑制剂阻断,而非程序性细胞死亡则是细胞被动的死亡过程,不能被细胞信号转导的抑制剂阻断。
非程序性细胞死亡即坏死,是指细胞在受到环境中的物理或化学刺激时所发生的细胞被动死亡[3]。其主要形态学特点是胞膜的破坏,细胞及细胞器水肿。
8(1)综述:细胞死亡的分类
细胞死亡后,细胞内容物及前炎症因子释放,趋化炎症细胞浸润引起炎症,以去除有害因素及坏死细胞并进行组织重建。哈佛大学医学院的袁均英教授和她的研究小组通过高通量的筛选,找到了一种化学小分子Necrostatin-1,能够抑制Fas/TNFR引起的坏死。该分子能专一性地阻断细胞坏死,但对凋亡没有抑制作用,这种死亡方式被他们命名为Necroptosis。他们的研究还指出,坏死也能由细胞信号通路介导。所以说,坏死不都是非程序性的,很多信号通路介导坏死的发生,其原因可能是由于认识的局限性,很多和坏死形态相似的死亡被误认为是坏死,因此应该把传统意义的坏死分为两类,即坏死和坏死样程序性细胞死亡。

三、程序性细胞死亡
程序性细胞死亡的分类方式大致有两种,即基于死亡机制的分类和基于形态学的分类。基于机制可以将程序性细胞死亡分为两大类:Caspase依赖的和Caspase非依赖的。前者即典型的凋亡,后者包括自吞噬性程序性细胞死亡、Paraptosis、细胞有丝分裂灾难、胀亡等。基于形态学的分类又有两种:一种将程序性细胞死亡分为凋亡、凋亡样程序性细胞死亡和坏死样程序性细胞死亡;另一种则分为I类、II类和III类程序性细胞死亡。
(一)基于机制的程序性细胞死亡分类
1.凋亡
1972年Kerr从形态学的角度描述了细胞的生理死亡,并将其命名为凋亡。凋亡的形态学特征为染色质凝集、边缘化,细胞皱缩,细胞膜内侧的磷脂酰丝氨酸外翻,细胞出泡形成凋亡小体。细胞凋亡受到一系列相关基因的严格调控。根据凋亡信号的来源可以将细胞凋亡信号转导通路分成两条:外源性通路(死亡受体通路)和内源性通路(线粒体通路)。两条信号通路汇集于下游的效应Caspase。效应Caspase在细胞凋亡的执行阶段能够直接引起重要蛋白质的降解和核酸酶的激活并最终导致细胞凋亡。也有一些凋亡不依赖于Caspase,而是通过AIF等分子发挥效应,其形态学变化属于凋亡样程序性细胞死亡。
2.自吞噬性程序性细胞死亡
1966年Duve和Wattiaux在发现溶酶体的同时发现了细胞的自吞噬现象[8],1977年Mortimore和Schworer发现肝细胞在处于饥饿状态时,自吞噬对其维持自身的稳态发挥着至关重要的作用。自吞噬细胞形态学上最主要的特征是细胞内出现大量泡状结构,即双层膜自吞噬泡,吞噬泡内为胞质及细胞器。调控自吞噬的细胞转导信号有很多,其中相对比较清楚的是PI3K和mTOR。PI3K对于早期吞噬泡的形成至关重要,而mTOR则对吞噬泡的形成及成熟起抑制作用。

8(2)综述:细胞死亡的分类
外已知有十数种ATG分子参与了吞噬泡的形成。当细胞处于氨基酸饥饿、营养缺乏或生长因子去除时,细胞的mTOR就会受到抑制,从而发生自吞噬。自吞噬是细胞在处于恶劣环境时的一种生存机制,但持续的自吞噬会导致程序性细胞死亡。3.Paraptosis 2000年Sperandio等在293T细胞系中超表达胰岛素样生长因子1受体(IGFIR)时发现一种与经典凋亡不同的死亡表型,并定义为Paraptosis。Paraptosis的形态学特征是细胞浆空泡化,线粒体和内质网肿胀,但没有核固缩现象。现在Paraptosis的文献报道比较少,其机制有待于进一步深入研究。高剂量的IGF或胰岛素等营养因子可以通过IGFIR活化MAPK/ERK以及JNK通路引起Paraptosis的发生,并且可以被AIP1/Alix特异性地抑制。我们实验室的王莺博士发现TNF受体超家族成员TAJ/TROY也可以诱导Paraptosis,并且可以被PDCD5加强[。
4.细胞有丝分裂灾难
1989年,Lisa Molz等发现在酵母的一种对热敏感的突变株中,细胞分裂时染色体分离发生异常。相应的一些学者便把这种在DNA发生损害时,细胞无法进行完全的分裂从而导致四倍体或多倍体的现象称为细胞有丝分裂灾难。对于细胞有死分裂灾难的形态学特点描述并不是很完全,但主要是巨细胞的形成,内有多个小核,染色质凝聚。有丝分裂灾难和凋亡的染色体固缩是否一样,现在看法并不一致。DNA发生损害时,如果细胞不能有效地阻断其细胞周期的进行,会导致染色体的异常分离,这些非正常分裂的细胞在下一轮有丝分裂中会继续导致细胞多倍体的形成从而成为癌变的基础。而细胞有丝分裂灾难作为一种死亡机制可以使这种非正常分裂的细胞死亡。细胞有丝分裂灾难由多种分子调控,如CDK1,P53及Survivin等,其死亡信号传递有很大一部分与凋亡相重叠[。
5.胀亡
1910年,von Reckling-hausen在骨软化病中发现由于缺血而肿胀坏死的骨细胞,他把这种肿胀坏死叫做Oncosis。1995年Majno和Joris为与凋亡相区别,重新引入了Oncosis的概念,把具有明显肿胀特点的细胞死亡命名为Oncosis,中文一般翻译为胀亡。胀亡的形态学特征是细胞肿胀,体积增大,胞浆空泡化,肿胀波及细胞核、内质网、线粒体等胞内结构,胞膜起泡,细胞膜完整性破坏。胀亡细胞周围有明显炎症反应。对于胀亡发生的机制现有的文献阐述较少,有研究者认为胀亡只是坏死前的一个被动性死亡阶段。但是近年来的研究更倾向于胀亡是一个程序性的死亡方式,如抗Porimin的单抗可以引起Jurkat细胞的胀亡;解偶联蛋白高表达的组织或细胞易发生胀亡,其原因是这些细胞缺乏ATP,故其膜上的离子泵功能丧失从而导致胀亡,而抑制解偶联蛋白可以抵抗胀亡却不能抵抗凋亡,提示胀亡可能有与凋亡不同的死亡机制。纽荷尔细胞显微镜使用方法。
8(3)综述:细胞死亡的分类
(二)基于形态的程序性细胞死亡分类
1.基于细胞核改变的分类
细胞核在细胞死亡时变化比较明显,所以很多人以此为标准将细胞死亡分为凋亡、凋亡样程序性细胞死亡、坏死样程序性细胞死亡和坏死,其中前三种是程序性细胞死亡,坏死是非程序性细胞死亡。
如前所述,凋亡细胞核的特点是染色质凝聚,成球状或半月状。其它的形态学变化还有磷脂酰丝氨酸外翻、细胞皱缩、凋亡小体形成等。凋亡一般都会伴有Caspase尤其是Caspase3的活化。
凋亡样程序性细胞死亡核的特征是染色质凝聚的程度较低,比凋亡细胞的染色体疏松一些,同时可以有凋亡细胞其它方面的形态学的变化,多数文献中描述的Caspase非依赖的凋亡归于此类。
坏死样程序性细胞死亡一般无染色质的凝聚或者只有疏松的点状分布,一些特殊的Caspase非依赖的死亡归于此类。
线粒体在细胞死亡中扮演着核心角色,其所释放的促进细胞死亡的分子大致有三类。第一类是细胞色素C,它与胞浆里的Apaf-1,ATP和Caspase-9前体分子形成Apoptosome,引起Caspase-9的激活,进而激活Caspase-3,引起典型的凋亡。第二类是超氧自由基,可以使细胞发生坏死样程序性细胞死亡,且此效应剂可以被抗氧化剂所拮抗。第三类是AIF,AIF可以切割染色体产生大的DNA片段(约50kB),而Caspase激活的DNA酶切割DNA产生的片段则很小(几十个碱基对),二者对细胞核形态的影响也不相同。AIF可以使细胞发生凋亡样程序性细胞死亡[24]。
2.Clarke分类
1990年Clarke等补充了Schweichel和Merker对于细胞死亡的分类,将程序性细胞死亡分为I类、II类和III类,至今仍被广泛引用。
I类程序性细胞死亡即是凋亡,形态学特征如上所述,这类死亡一般没有溶酶体的参与,且死后会被吞噬细胞所吞噬。

II类程序性细胞死亡即是自吞噬性程序性死亡,如上所述,其主要的形态学特征是自吞噬泡的形成,自吞噬泡和溶酶体融合后被消化,而细胞残骸会被吞噬细胞吞噬。
III类程序性细胞死亡即是坏死样程序性细胞死亡,其主要的形态学特征是各种细胞器的肿胀、胞膜的破坏等,这类细胞死亡没有溶酶体的参与。III类程序性细胞死亡又分为两个亚类IIIA和IIIB,其中IIIB亚类胞膜破坏比较轻微,各类细胞器的肿胀表现比较明显,而且死亡后会被吞噬细胞吞噬。
8(4)综述:细胞死亡的分类
上述两种分类方法都涉及坏死性程序性细胞死亡,但是两种方法所关注的形态学特征却并不一致。Clarke分类中的II类程序性细胞死亡如果按照细胞核形态分类应该属于坏死样程序性细胞死亡,因为它没有发生染色质凝聚。
(三)形态学分类与机制分类的关系
上述程序性细胞死亡分类似乎有些复杂,因为形态学分类与机制分类具有很大的重叠;所以,如果能够将二者联系起来,找出它们的对应关系,将有利于我们对于细胞死亡的理解。
1、机制分类中的凋亡按核形态分类属于凋亡或凋亡样程序性细胞死亡;Clarke分类属于I型程序性细胞死亡。
2、机制分类中的自吞噬性程序性细胞死亡按核形态分类应该属于坏死样程序性细胞死亡,因为其没有明显的染色质凝聚;Clarke分类属于II型程序性细胞死亡。
值得注意的是,凋亡可以以自吞噬开始,而自吞噬可以以凋亡结束,其机制为溶酶体的活化可以激活典型的内源性凋亡通路。所以,很多自吞噬性程序性细胞死亡结果表现为凋亡,但也有文献报道有独立的自吞噬性程序性细胞死亡,同时又没有凋亡发生。
3、机制分类的Paraptosis按核形态分类应该属于坏死样程序性细胞死亡,纽荷尔细胞显微镜使用方法,因其核没有明显的变化;Clarke分类属于III型程序性细胞死亡。
4、机制分类的细胞有丝分裂灾难严格来说应该归于凋亡,因为其表现出明显的染色体凝聚。但由于其诱因比较特殊,所以习惯上把它单独归于一类。
5、机制分类的胀亡按核形态分类应该属于坏死样程序性细胞死亡,因为其染色体可以形成散在的点状分布[31];Clarke分类属于III型程序性细胞死亡。
随着近年来对于细胞死亡的深入研究,经典的“凋亡——坏死”的死亡分类模型已经显得过于简单。一般来说,某一种死亡刺激可能引起细胞多个死亡通路的活化,细胞最终发生什么样的死亡主要取决于被活化通路发挥作用的速度。多数情况下,Caspase通路发挥作用最快,因此程序性细胞死亡最常表现为凋亡。但是某些情况下,比如Caspase通路受阻时,Caspase通路不发挥作用或者速度慢于其它通路,细胞就会表现为凋亡样或坏死样程序性细胞死亡。在某些Bcl-2高表达的肿瘤细胞系中,死亡刺激也往往引起凋亡样或坏死样程序性细胞死亡。细胞具有多种死亡方式是有其进化必然性的。多细胞生物在生命过程中会有大量“多余的”或“生病的”细胞死掉,单一的死亡方式对于生命来说是十分危险的,因为一旦此通路受到抑制,后果将不可想象。多种死亡方式的存在就像一个多重保险,会“千方百计”地使应该死亡的细胞死掉,即便是某些死亡通路受阻。现在对于细胞死亡的分类还有一些混乱,随着对于细胞死亡机制认识的不断深入,我们期待着一个更为合理的细胞死亡分类方法。



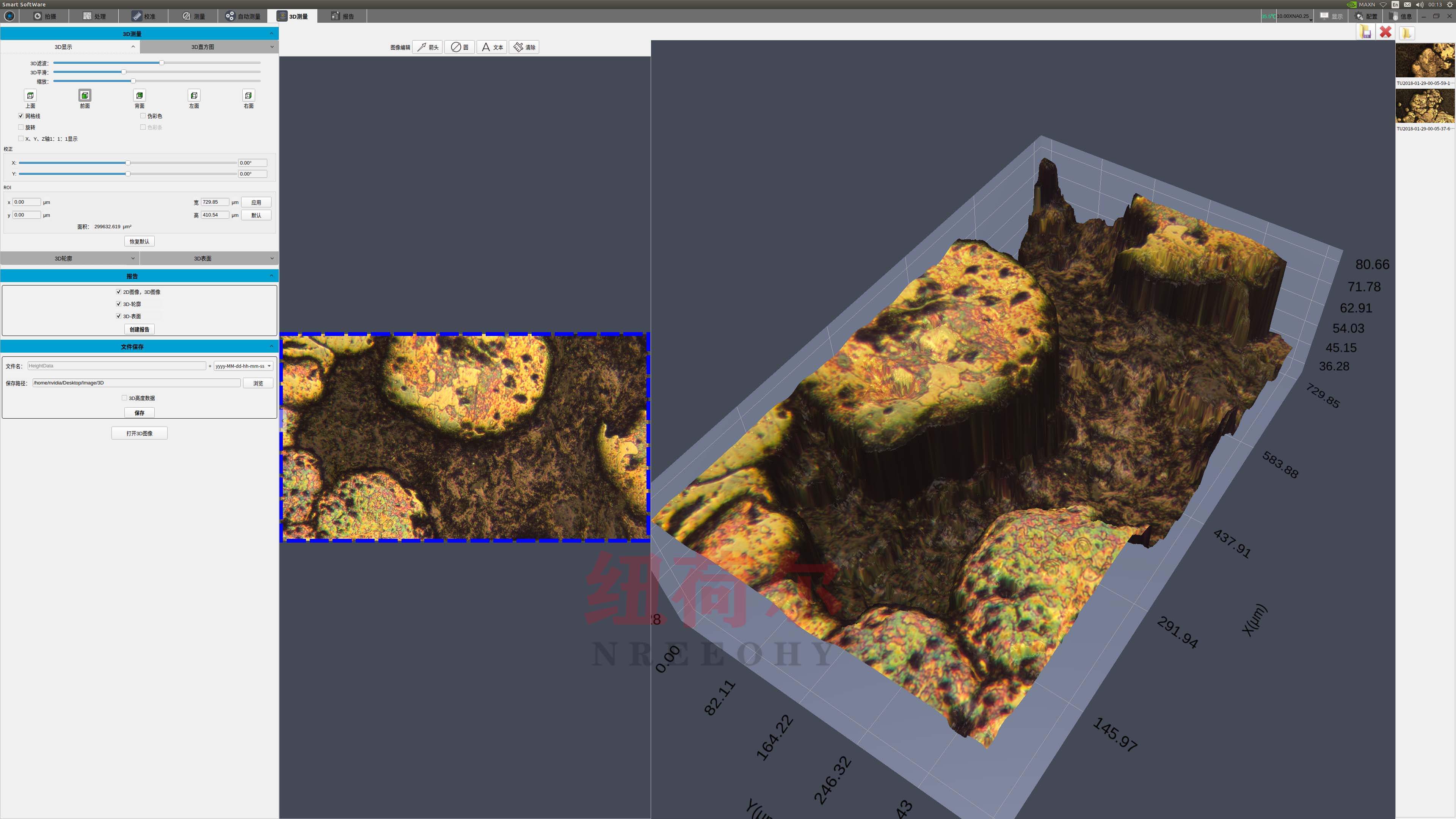
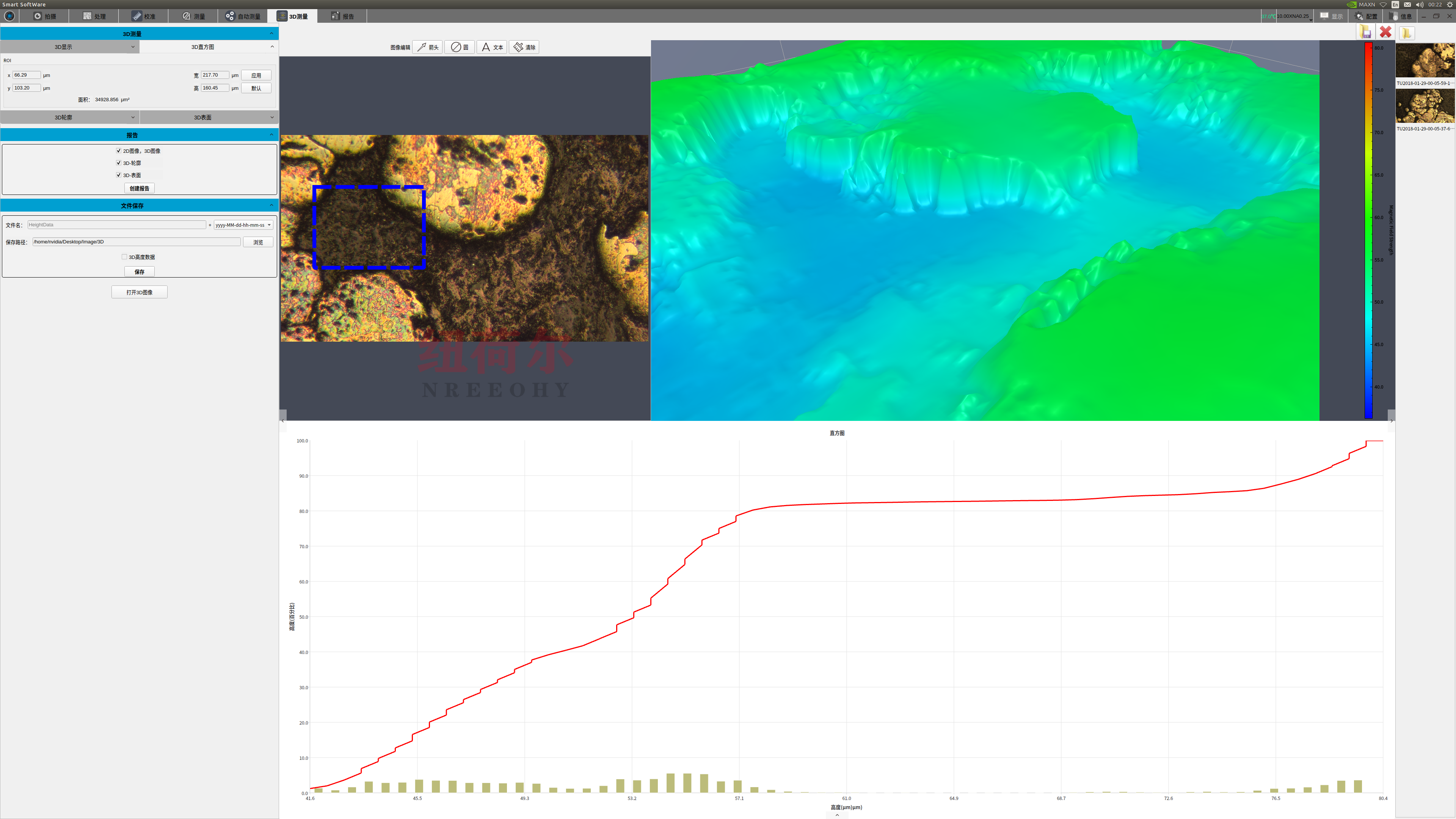
我们拥有的3D形状扫描测量显微镜,为全球客户提高质量、效率和生产率。
© 版权所有 深圳纽荷尔科技有限公司

您要复制的内容
采购电话
联系任一团队,我们很乐意为您提供指导


经销商中心
更多经销商中心提供最新的营销和销售支持材料。
微信公众号
扫码关注微信 了解更多资讯
支持
常见问题


实用链接
网站链接
新闻资讯
最新活动
客户评价
产品资料
政策资质
公司
关于我们
工作机会

